
Métabolisme et résistance à l'aridité
Michel Cornet
Parution in Succulentes 1990, n°1 (pp 3 - 6), n°2 (pp 3 - 10)
1- Une affaire d'enzymes
L'adaptation des plantes à l'aridité revêt de nombreux aspects que révèle une observation attentive. Mais le métabolisme, dont les étapes ne sont connues que par des expériences délicates, n'est pas moins important pour cette adaptation et ses caractéristiques chez les plantes des régions arides sont pleines d'enseignements (1).
Toutes les plantes chlorophylliennes utilisent la lumière comme source d'énergie pour construire, à partir du bioxyde de carbone (CO2) de l’atmosphère, et de l'eau, les substances carbonées (Glucides...) qui leur sont nécessaires : c'est la photosynthèse, classée en trois types de métabolisme.
La figure 1 rappelle les 3 modes d'incorporation du CO2 (carboxylation) et souligne les caractères communs et les différences des trois métabolismes. Le même cycle photoréductif (cycle de Calvin) intervient pour toutes les plantes, mais la carboxylation initiale est différente pour les végétaux de type C3 d'une part (le CO2 atmosphérique entre directement dans le cycle de Calvin) et les végétaux de type C4 ou CAM d'autre part (le CO2 atmosphérique est incorporé au cours d'un premier cycle précédent le cycle de Calvin). Les végétaux de type CAM (Métabolisme Acide Crassulacéen) ont en outre un avantage majeur pour vivre en régions arides : l'incorporation du CO2 est nocturne, ce qui permet la fermeture des stomates le jour et diminue considérablement les pertes d'eau par transpiration lorsque la température foliaire est la plus élevée (1).
Dans les cycles métaboliques, à diverses étapes interviennent des enzymes, protéines catalysant des réactions chimiques, impossibles sans elles aux températures compatibles avec la vie. Leur rôle est essentiel. La Phosphoénol Pyruvate Carboxylase (PEPCase), qui catalyse la carboxylation, la nuit chez les plantes de type CAM, le jour chez les plantes de type C4, est beaucoup plus active (4 à 10 fois plus rapide (2)) que la Ribulose Bisphosphate Carboxylase (RuBPCase, la protéine la plus répandue dans le monde vivant), qui catalyse, à la lumière, la carboxylation chez toutes les autres plantes, de type C3.
Cette grande activité de la PEPCase a une importante conséquence : pour une même quantité d'eau utilisée par l'organisme, les plantes de type C4 ou CAM des régions sèches ou arides ont un pouvoir de fixation du CO2, donc de croissance, plus élevé que les plantes de type C3 des régions tempérées humides (3). Plus précisément, le rendement énergétique de leur photosynthèse est doublé par rapport à celui des plantes C3 (2), avec un taux de transpiration pour les plantes de type CAM deux fois plus faible que pour les plantes C4 et quatre fois plus faible que pour le plantes C3 (3). Chez les plantes C4 et CAM, l'incorporation du CO2 n'est pas limitée par la faible concentration de ce gaz dans l'atmosphère (0,03 pour cent), comme c'est le cas pour les plantes C3, mais par l'intensité lumineuse. C'est un avantage dans les milieux où cette intensité est élevée. Au contraire, l'activité photosynthétique des plantes C3 plafonne dès 25 % environ de l'ensoleillement maximal (2). Chez les plantes C3, la RuBPCase catalyse, à la lumière, non seulement la carboxylation de la RuBP, mais aussi son oxydation par incorporation d'oxygène : ce phénomène parallèle de photorespiration se traduit par le rejet de CO2, donc par une diminution importante du rendement photosynthétique, de 30 à 50 % (2), et cela d'autant plus que la température est élevée. Au contraire, l'activité de la PEPCase peut fournir, chez les plantes C4 ou CAM, des quantités importantes de CO2 qui, en bloquant l'activité oxydase de la RuBPCase, bloquent la photorespiration quelle que soit la température. Enfin, la faible humidité provoque une fermeture partielle des stomates par lesquels s'effectuent les échanges gazeux dans tous les types de plantes. Cette fermeture partielle n'est pas un handicap pour la rapide PEPCase alors qu'elle le devient pour la RuBPCase, déjà lente pour une ouverture maximale des stomates.
Ainsi, les métabolismes C4 ou CAM permettent l'utilisation optimale des ressources d'un milieu sec ou aride caractérisé par de grandes luminosités, des températures élevées, une faible humidité, et ils ne sont pas limités par la faible concentration de CO2 dans l'atmosphère. Les plantes de type C4 ont été présentées comme des «crypto-succulentes» (4) ; le type CAM est, sauf exceptions, particulier aux plantes à morphologie et anatomie succulentes.
En effet, le type CAM possède en propre des avantages en situation très aride. Par rapport au métabolisme C4, il se caractérise par des mécanismes de régulation et d'adaptation qui favorisent une plus grande résistance à de longues sécheresses. Le premier intervient au cours du cycle journalier. Un signal «début de nuit» déclenche l'activité de la PEPCase, la production des corps à quatre atomes de carbone (Malate et/ou Aspartate) et leur stockage. Ces produits étant des inhibiteurs de la PEPCase, leur accumulation dans les vacuoles foliaires ralentit, puis bloque l'entrée de CO2. Un second signal «début de jour» permet ensuite la décarboxylation des corps à quatre atomes de carbone et l'utilisation par le cycle photoréductif du CO2 produit (5). L'enzyme catalysant la décarboxylation est en effet inhibée par les faibles températures nocturnes et favorisée par l'élévation de température diurne (3). Ce premier mécanisme régulateur est donc essentiel pour que la plante puisse absorber le CO2 la nuit, avec un minimum de perte d'eau par transpiration. Il fonctionne pour des températures nocturnes relativement basses ; par contre, si la température nocturne s'élève, la fixation de CO2 pendant la nuit diminue et la fixation diurne prend plus d'importance (cas d'Agave americana (3,6)).
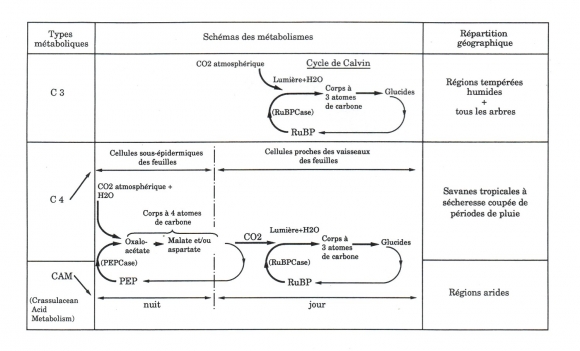
Figure 1 - Les trois types de métabolisme des plantes chlorophylliennes. RuBP : Ribulose biphosphate. (RuBPCase) : Ribulose biphosphate carboxylase, catalyseur de la carboxylation de la RuBP. PEP : Phosphoénol pyruvate. (PEPCase) : Phosphoénol pyruvate carboxylase, catalyseur de la carboxylation du PEP.
Certaines espèces ne sont pas de type CAM en permanence. Sous l'effet d'une augmentation de température ou d'une période de sécheresse, elles modifient profondément leur métabolisme et adoptent le type CAM. La salinité du sol peut également intervenir (6). Chez d'autres espèces, c'est la photopériode (durées relatives du jour et de la nuit) qui déclenche le type CAM. Kalanchoe blossfeldiana, K. velutina, K. daigremontiana, K. tubiflora sont dans ce cas. En jours longs, l'activité de la PEPCase est maintenue à un faible niveau : la principale voie d'entrée du CO2 est alors catalysée par la RuBPCase (comportement de type C3). En jours courts, l'augmentation d'activité de la PEPCase est exponentielle : il en résulte un comportement de type CAM (5). La transpiration diurne est réduite de trois fois par rapport aux échanges de jours longs. Ainsi, le photopériodisme assure une préparation de la plante à une contrainte hautement probable dans la saison à venir (7).
La photopériode possède aussi une grande influence sur l'activité de la PEPCase chez des espèces de type CAM permanent comme Opuntia ficus-indica (7). Lorsque cette espèce est soumise à une sécheresse modérée, le CAM est exalté. Dans le cas d'une sécheresse extrême (jours longs, thermopériode 40/30°C, pas d'arrosage), les stomates se ferment et l'absorption du CO2 atmosphérique cesse. Cependant, la PEPCase, en reprenant le CO2 interne produit par la photorespiration, continue à maintenir le métabolisme CAM en activité, bien qu'à un niveau faible. Ce comportement, appelé CAM-idling, peut persister longtemps sans dommage : un mois quel que soit le traitement photopériodique préalable, jusqu'à 70 jours après une période de jours courts. Un arrosage fait reprendre aussitôt l'absorption nocturne de CO2. Dans ce cas encore, une période de jours courts accroît donc la résistance ultérieure à la sécheresse intense, situation très probable au cours des jours longs suivants.
Par leur économie hydrique très efficace, les plantes de métabolisme C4 ou CAM possèdent donc des avantages incontestables en situation sèche. Cependant, les deux solutions ne sont pas équivalentes (6). Les plantes de type C4, principalement originaires des régions tropicales, peuvent résister à la sécheresse puis croître très rapidement lorsque l'eau devient disponible : le type C4, aidé par une anatomie foliaire particulière (4), est une optimisation du métabolisme en présence de forts éclairements (6). Les plantes de type CAM, principalement des succulentes pour lesquelles le faible rapport de la surface externe au volume minimise la transpiration, mais également les échanges gazeux, n'ont pas ces possibilités de croissance. Mais elles sont mieux adaptées aux milieux dans lesquels la pression de sélection provient essentiellement de l'utilisation de l'eau (4).
2- Un exemple de convergence évolutive
Selon les espèces et les conditions du milieu, le Métabolisme Acide Crassulacéen (CAM) se présente, comme nous l'avons vu précédemment, sous des degrés divers. Certaines plantes succulentes adoptent le type CAM sous l'effet de contraintes extérieures (sécheresse, salinité du sol, température, photopériode), d'autres sont de type CAM en permanence, d'autres, telles Portulacaria afra ou Senecio articulatus sont mixtes, de type CAM la nuit et C3 le jour (3). D'autres encore (Sedum telephium (8)) sont mixtes C3-CAM et deviennent de type purement CAM sous l'effet d'un manque d'eau. Toutes les plantes C3, C4 ou CAM possèdent en effet l'enzyme nécessaire au métabolisme CAM (comme au C4), la Phosphoénol Pyruvate Carboxylase (PEPCase), mais cette dernière n'a qu'une activité très faible chez les plantes de type C3, quinze à vingt fois inférieure à son activité dans le type C4 par exemple (3, 6) : on désigne ces enzymes de compositions semblables mais de propriétés différentes sous le nom d'isoenzymes. Le mécanisme de passage de la forme peu active à la forme active ne semble pas encore connu, mais on sait qu'il peut s'accompagner de transformations anatomiques (8).
Tableau 1 : Liste des familles comportant des espèces de type C4 ou CAM, ou des Succulentes.
Légende tableau ci-dessus.
- Classification des Angiospermes selon J.-F. Leroy (9), des plantes autres que les Angiospermes selon A. Takhtajan, «Floristic Regions of the World», University of California Press, USA, 1986.
- Liste des plantes C4 : M.-L. Champigny et A. Moyse (6) d'après A.-S. Raghavendra et V.S.R. Das,Photosynthetica, 12 (1978) p. 200-208. - Liste des plantes CAM : M.-L. Champigny et A. Moyse (6) d'après S.-R. Szarek, Photosynthetica, 13 (1979) p. 467-473.
- Liste des plantes succulentes établie d'après H. Jacobsen, «A Handbook of Succulent Plants», Blandford, 1986. Cactaceae d'après A. Takhtajan.
(Les chiffres de la colonne «Succ» indiquent, le premier le nombre de genres possédant des espèces succulentes, le second le nombre total d'espèces succulentes dans la famille).

Ces nombreux comportements intermédiaires, l'existence des isoenzymes, semblent indiquer le résultat d'une évolution en réponse à des contraintes du milieu. W.M. Laetsch (4) a émis l'hypothèse, reprise ensuite (3), d'une adaptation à la présence, en quantité importante, de cations (c'est-à-dire à l'existence d'une salinité) dans les sols des régions arides pour expliquer l'apparition des métabolismes C4 et CAM. Le cation Na+ est en effet nécessaire aux espèces de type C4 ; il révèle également la tendance CAM de deux Mésembryanthémacées succulentes, par ailleurs de type C3, M. cristallinum et Aptenia cordifolia (3, 6).

Quel qu'en soit le mécanisme, l'évolution de certaines familles de plantes pour aboutir aux types C4 ou CAM s'est probablement effectuée relativement tard dans l'évolution végétale. En effet, la liste des familles comprenant des espèces de type CAM ou C4, dressée selon une classification phylogénétique moderne exposée par J.-F. Leroy (9), comporte (tableau 1) une Isoëtacée (8), au moins deux Polypodia-cées (6), ainsi que Welwitschia, espèce succulente de niveau de développement intermédiaire entre celui des Gymnospermes et celui des Angiospermes (ou plantes à fleurs), mais la très grosse majorité des plantes CAM ou C4 se trouve parmi les Angiospermes. De plus, peu d'espèces appartiennent aux groupes considérés comme les plus primitifs parmi les Angiospermes : la sous-classe des Magnoliidées chez les Dicotylédones et, dans une moindre mesure, celle des Alismatidées chez les Monocotylédones. Les métabolismes CAM ou C4 apparaissent dans toutes les autres sous-classes excepté celle des Hamamélididées, dont beaucoup d'espèces sont encore archaïques (9) et celle des Palmiers (Arécidées).


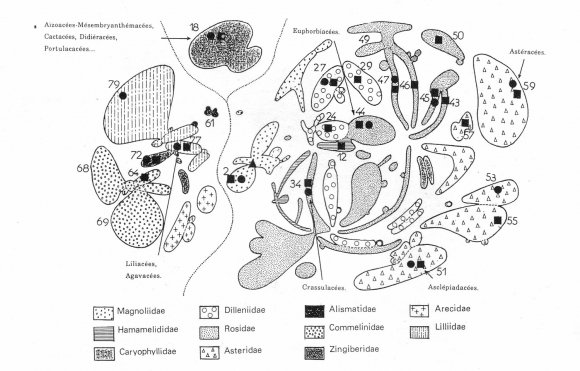
Les numéros renvoient aux ordres du tableau 1.
● Métabolisme CAM. ■ Espèces succulentes.
▲ Noyau primitif des Angiospermes, proche des Chloranthacées.

Les familles comprenant des espèces succulentes se trouvent de même dispersées dans toutes les sous-classes, les plus primitives exceptées. Les grandes familles de plantes succulentes illustrent cette dispersion. Parmi les Rosidées se placent les Euphorbiacées avec leur organisation florale en cyathium, signe d'évolution (9), et les Crassulacées. Les Asclépiadacées au système de pollinisation très élaboré, semblable à celui des Orchidacées, appartiennent aux Astéridées, une sous-classe très évoluée que couronnent les Astéracées, dont Senecio est l'un des principaux genres. Chez les Monocotylédones, les Liliacées, comme les Agavacées qui en dérivent (10), appartiennent à l'ordre des Liliales. L'ordre des Caryophyllales (Dicotylédones) qui caractérise la sous-classe des Caryophyllidées est la plus riche en espèces succulentes : Aïzoacées-Mésembryanthémacées, Cactacées, Didiéracées, Portulacacées... Les trois premières familles sont proches les unes des autres et les plus évoluées de la sous-classe (10). Dans cet ordre très homogène des Caryophyllales se rencontrent également 85 % des plantes à métabolisme C4 (9).
Ces degrés d'évolution sont résumés dans la figure 2, coupe horizontale de l'arbre «généalogique» probable des plantes à fleurs. Cette coupe, établie selon J.-F. Leroy (9) d'après des travaux de Dahlgren (1975) et de Cronquist (1978), présente la situation actuelle des Angiospermes. Les ordres apparaissent sous forme de bulles en raison du nombre de leurs espèces ; ils sont disposés selon leurs relations de parenté établies en considérant le plus grand nombre disponible de caractères. Les ordres sont d'autant plus évolués qu'ils s'écartent plus du noyau primitif des Angiospermes, probablement très proche des Chloranthacées (11, 12, 13) (Magnoliidées). Les grandes familles de plantes succulentes occupent des emplacements très divers, éloignés les uns des autres : leurs évolutions respectives ont été très différentes. Les familles comportant des espèces succulentes (marquées ■ sur la figure 2) se répartissent sur tout le diagramme, souvent à la périphérie, indiquant qu'une évolution importante a eu lieu par rapport au type ancestral (marqué ▲). À la succulence est souvent associée la présence du métabolisme CAM (marqué ●) : sur 44 familles comprenant des espèces succulentes, 20 possèdent également des espèces CAM.
En conclusion, le Métabolisme Acide Crassulacéen est apparu en relation étroite avec la succulence, dans des familles de plantes très éloignées les unes des autres par leurs origines. Ce métabolisme est le résultat «d'une convergence fonctionnelle évolutive» (6) dont les formes intermédiaires signalées au début pourraient représenter des réalisations inachevées. La position de ces familles dans l'arbre phylogénétique traduit le plus souvent, parmi les Angiospermes, un degré d'évolution important qui n'est pas seulement le reflet de l'acquisition de la succulence et du Métabolisme Acide Crassulacéen.
BIBLIOGRAPHIE
(1) C. BULARD, Succulentes (1985) n° 1, p. 16.
(2) C.R. SOMERVILLE et S.C. SOMERVILLE, La Recherche, 154 (1984), p. 490-501.
(3) A. MOYSE, Physiolol. vég., 14 (1976) p. 533-550.
(4) W.M. LAETSCH, Ann. Rev. Plant Physiol., 25 (1974) p. 27-52.
(5) O. QUEIROZ, Physiol. vég., 14 (1976) p. 629-639.
(6) M.-L. CHAMPIGNY et A. MOYSE, Bull. Soc. bot. Fr., 130, Actual. bot., 1 (1983) p. 71-83.
(7) J. BRULFERT, D. GUERRIER et O. QUEIROZ, Bull. Soc. bot. Fr., 131, Actuel, bot., 1 (1984) p. 69-77.
(8) P.-D. MOORE, Nature, 337 (1989) p. 694.
(9) J.-F. LEROY, «Les Angiospermes» dans «Précis de Botanique-2 : Végétaux Supérieurs», Masson. Paris (1982), p. 201-473.
(10) A. CRONQUIST, «An Integrated System of Classification of Flowering Plants», Columbia University Press. New-York (1981).
(11) J.-F. LEROY, Taxon, 32 (2) (1983), p. 169-175.
(12) J.-F. LEROY, C.R. Acad. Soi. Paris, 296, série III (1983), p. 747-752.
(13) E.-M. FRIIS, P.-R. CRANE et K.-R. PEDERSEN, Nature, 320 (1986), p. 163-164.
Voir aussi / see also : Crassulacean Acid Metabolism CAM